
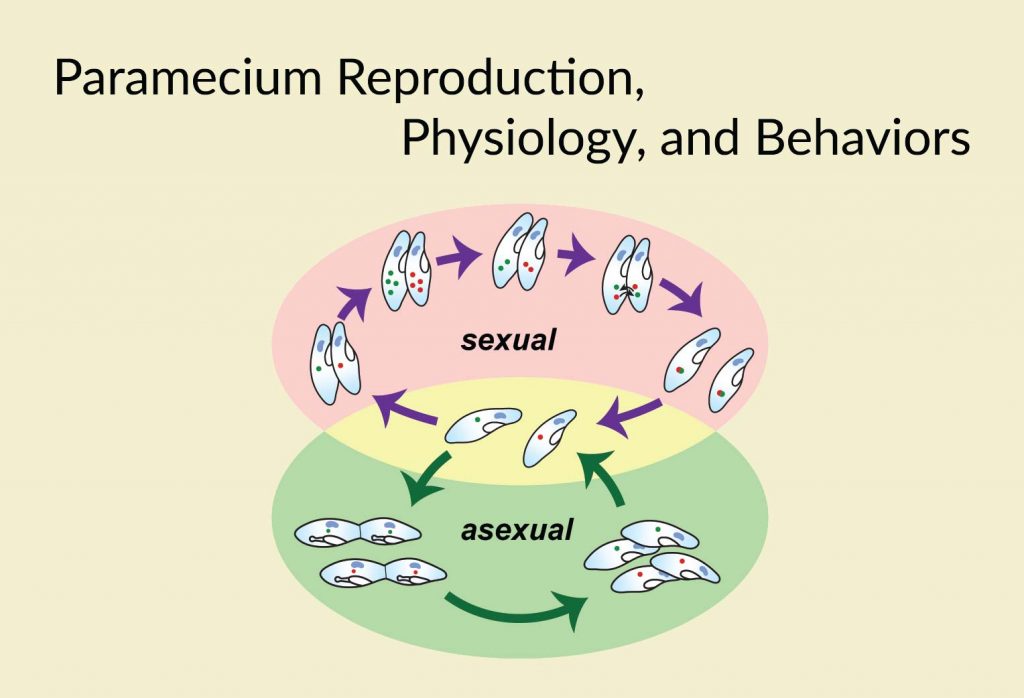
In this blog post, we will learn about paramecium reproduction, aging, learning and memory ability, movement, sensing, feeding behaviors, and their endosymbiotic relationship with algae.
We also have 4 series blog posts about paramecium:
Part I. The Biological Classification of Paramecium – Name, History, and Evolution
Part II. The Structure of Paramecium cell
Part III. Paramecium Reproduction, Physiology, and Behaviors
Part IV. The Natural Habitation and Cultivation of Paramecium – Find Paramecium for Your Microscopic Project
This article covers
How does a paramecium reproduce? – Life cycle of paramecia
Paramecia can reproduce either asexually or sexually, depending on their environmental conditions.
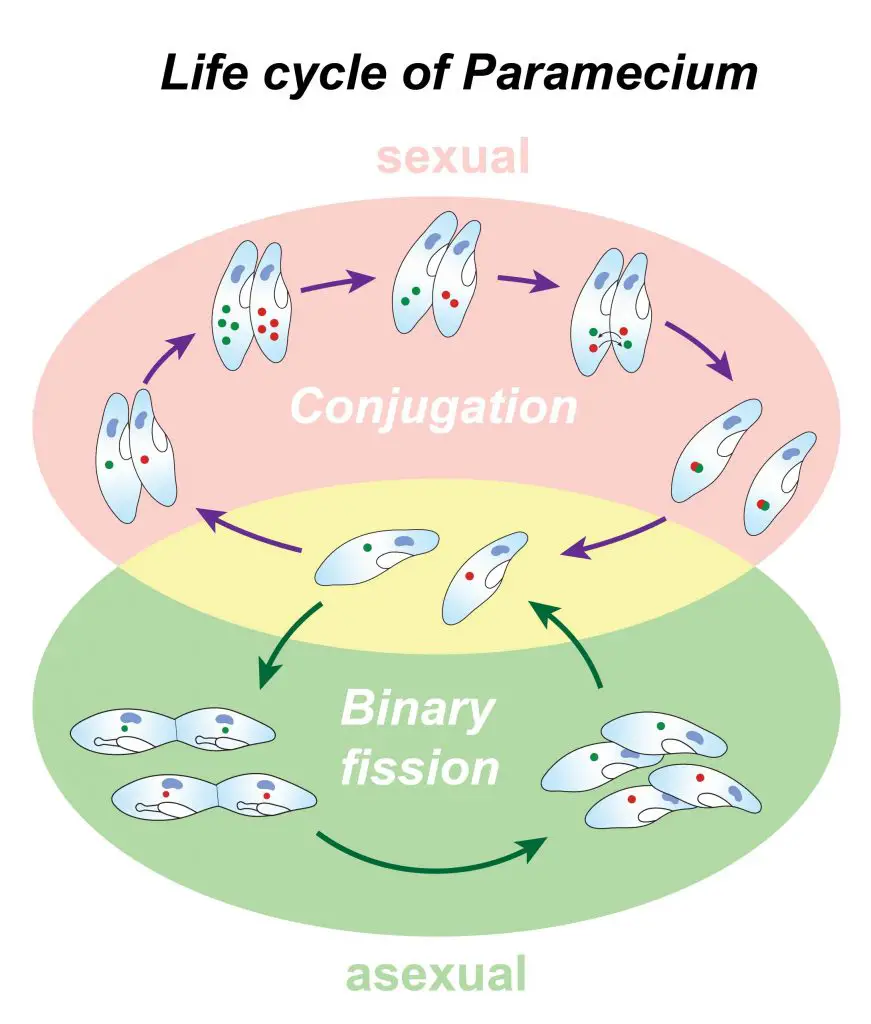
Asexual reproduction (binary fission)
Most of the time, paramecia reproduce asexually by splitting one cell into two cells, a process called “Binary Fission”. Binary Fission takes place when ample nutrients are available. Under favorable conditions, they may divide two or three times a day. The sizes of the paramecia population can grow rapidly by binary fission.
During binary fission, one paramecium cell divides into two daughter cells with identical genetical information. The micronucleus divides through “mitosis”, but the macronucleus divides another way, called “amitosis”.
Mitosis is the standard way of cell division that can guarantee the equal separation of parent cells’ chromosomes. Centrosomes project microtubule filaments to form a mitotic spindle and pull sister chromatids evenly toward two new nuclei.
The macronucleus can not undergo typical mitosis because it does not have a complete set of DNA and can not form paired sister chromatids. Instead, the macronucleus undergoes amitosis, which simply splits its DNA contents into two parts without spindle formation or the appearance of chromosomes. Interestingly, a new macronucleus after cell division somehow is able to keep approximately the same number of copies of each gene.
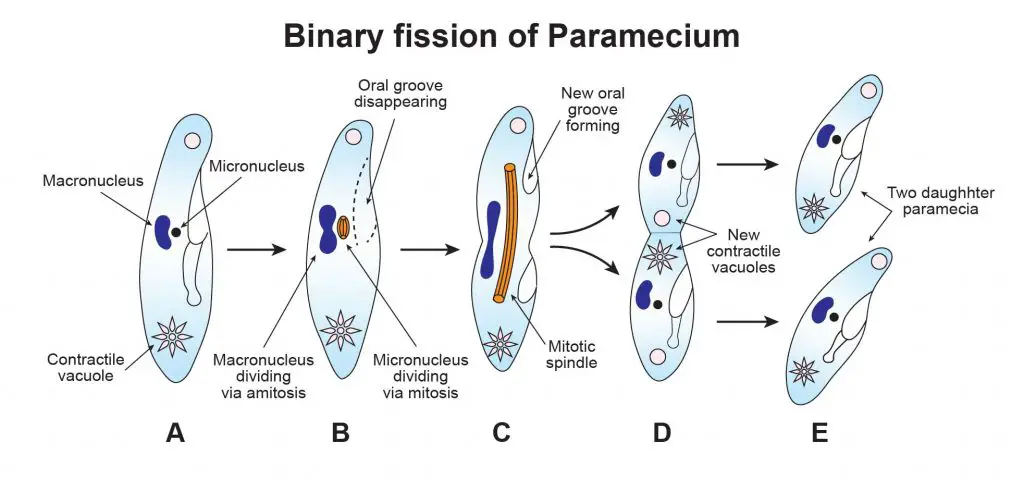
[In this figure] Binary fission of paramecium.
Paramecium reproduces asexually by binary fission during favorable conditions in the following steps: (A) Paramecium stops feeding and starts to replicate its DNA, (B) Oral groove disappears. The micronucleus forms the mitotic spindle and starts the mitotic division. The macronucleus changes its shape and starts the amitotic division. (C) Both mitosis and amitosis continue to split into two daughter micronuclei and macronuclei. New oral grooves form as the cell becomes elongated. (D) The endoplasm is divided into two in the middle of the cell transversely. The anterior portion is known as proter and the posterior portion is known as opishte. The proter and opisthe portions are just like that of their parents in size, shape, and structure. (E) The whole process completes. All such individuals which are formed from a single parent are known as a clone. One binary fission needs twenty minutes and in 24 hours it divides two or three times.
[In this video] Two Paramecium aurelia cells in the final stage of cell division.
At this stage (called cytogenesis), the division of nuclei has been completed. The rest of the cells (cytoplasm and organelles) divide to form two new cells.
Sexual reproduction (conjugation)
Sexual reproduction of paramecia takes place under conditions of starvation. There are two mating types for paramecia, which are referred to as odd and even. Odd types can only mate with the even types, but the same mating types can not mate with each other. Moreover, only cells within a single paramecium species can mate with one another (for example, P. aurelia and P. caudatum can not mate).
The process of sexual reproduction, also called conjugation, is easily distinguishable under the microscope. The mating cells stick together. In the laboratory, when two cultures of mating types are initially mixed, they actually form rather dramatic clumps of cells. Then those gradually pair off into individual pairs as the mating processes.
[In this video] A video showing two pairs of paramecia in conjugation.
The process of sexual reproduction (conjugation) is much more complicated than asexual reproduction (binary fission). Keep in mind: the reason why the paramecia decide to sexual reproduction is that they need to create “genetic variations” to increase their chance of survival under a harsh condition. In order to do so, two paramecia of compatible mating types exchange parts of their genetic materials and rearrange their DNA to create genetic variations. As a result, the offsprings of sexual reproduction have different genetic DNA sequences compared to their parents. On the other hand, the daughter cells generated from binary fission have identical genome like their parent cell. We call them “clones”.
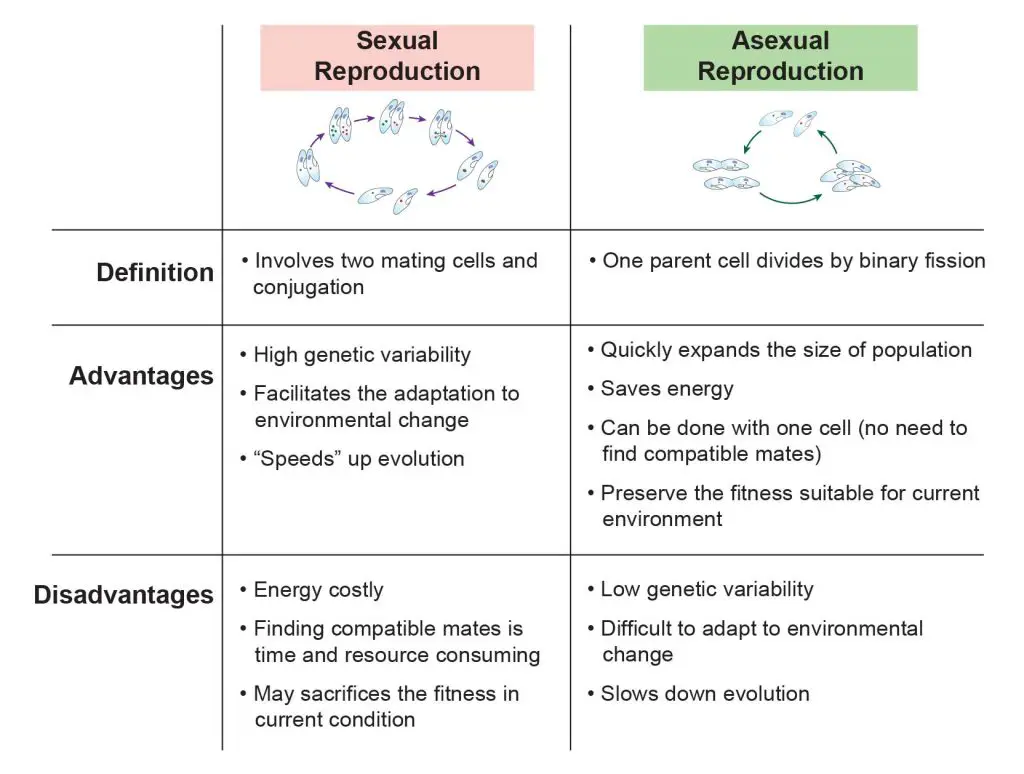
[In this figure] The comparison between sexual reproduction and asexual reproduction.
In brief, during conjugation of paramecium, the micronuclei of both paramecia undergo meiosis, ultimately halving the genetic content to create a haploid nucleus. The paired mating cells exchange one of their haploid nuclei. The haploid nuclei from each mate fuse to create a new, genetically varied, micronucleus. In turn, the new micronucleus replicates to give rise to a new macronucleus. For detailed step-by-step information on conjugation, see the schematic diagram below.
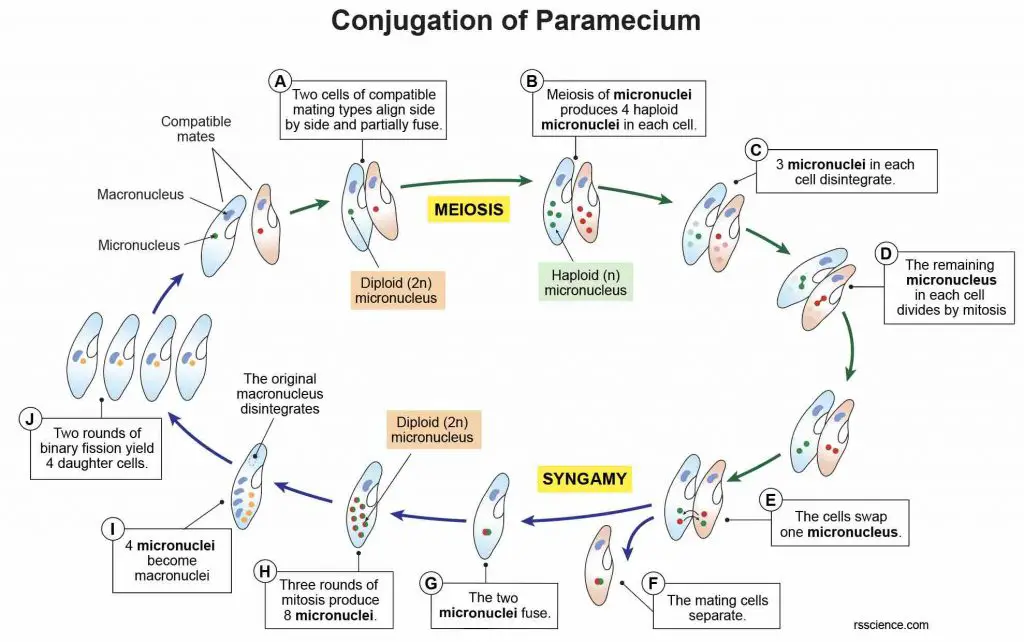
[In this figure] Steps of paramecium’s conjugation.
(A) Two cells of compatible mating types meet and align side by side. They become partially fused and form a protoplasmic bridge in-between. At this stage, their micronuclei are diploid (2n).
(B) Each cell undergoes meiosis to produce 4 haploid (n) micronuclei.
(C) Out of these 4 micronuclei, 3 degenerate and disappear.
(D) The remaining one micronucleus divides by mitosis to form two unequal pronuclei or gamete nuclei. The smaller gamete nucleus is active in migratory in nature and is called migratory gamete nucleus. The larger gamete nucleus is passive and stationary in nature and is called stationary gamete nucleus.
(E) The migratory gamete nucleus of one cell passes through the protoplasmic bridge into another cell.
(F) After the exchange of micronuclei, two mating paramecia separate.
(G) Within each cell, the new migratory gamete nucleus fuses with its stationary gamete nucleus. This fusion (also called syngamy) results in the formation of a single diploid zygote nucleus. This zygote nucleus is also known as synkaryon.
(H) The zygote nucleus in each cell divides three times by mitosis to form 8 nuclei.
(I) Four of these 8 nuclei grow in size to become macronuclei. The remaining 4 small nuclei are called new micronuclei.
(J) Out of the 4 micronuclei, 3 degenerate and disappear. And the remaining one micronucleus divides twice into 4 daughter paramecia, each has one micronucleus and one macronucleus. In this way, 8 paramecia are produced as a result of the conjugation of two mating paramecia.
Two significant advantages of conjugation for paramecium
Genetic variation
During conjugation, genetic materials are exchanged between the matching mating types. It brings genetic variability in the offspring individuals due to genetic recombination. Genetic variability can speed up evolution and increase the chance to adapt to environmental change.
Why does sexual reproduction increase the chance of survival of the entire population? See the illustration below.
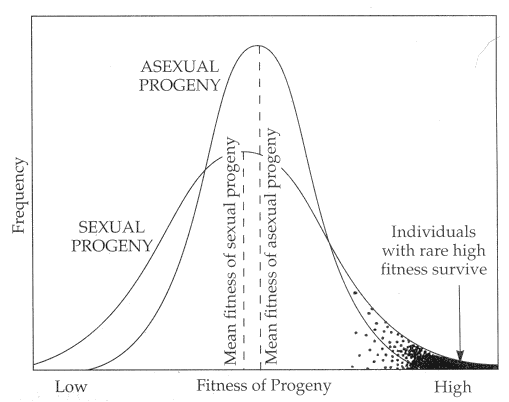
[In this figure] The fitness of sexual progeny vs asexual progeny.
The mean fitness of an asexual clone of organisms adapted optimally for a particularly narrow niche. They are perfect in the current environment, but, once the condition changes, the entire population may suddenly extinct. Sexual reproduction can produce a wider range of sub-optimally adapted types. Some may not be able to survive in the current environment (an example is genetic diseases in humans). However, an ecological disaster that eliminates the clonal niche will not have as severe an impact on the sexual clade, because of the diversity of genotypes.
Photo source: https://www.mun.ca/biology/scarr/Sexual_vs_Asexual_clades.htm
Rejuvenation
After conjugation, a new and metabolically active macronucleus is produced by a reorganization of micronuclear materials. The old, weak, and defective macronucleus is replaced by a new one. Rejuvenation can reset the aging after a prolonged clonal growth of asexual reproduction.
Autogamy (self-fertilization)
Under conditions of prolonged starvation, paramecia can also undergo “autogamy” or self-fertilization. Autogamy is essentially the same thing as conjugation, but it is only happening with a single cell.
During this process, the micronucleus replicates multiple times. One of these new micronuclei undergoes rearrangement of its genetic content. Some DNA is fragmented and some DNA sequences, known as “Internal Eliminated Sequences,” are removed. This can increase the genetic diversity to enhance the chance of survival in a harsh environment. As a result of autogamy, a new macronucleus is formed which rejuvenates the paramecium by increasing its vitality.
Autogamy is not unique to paramecium. Autogamy is frequently observed in many flowering plants as a form of self-pollination.
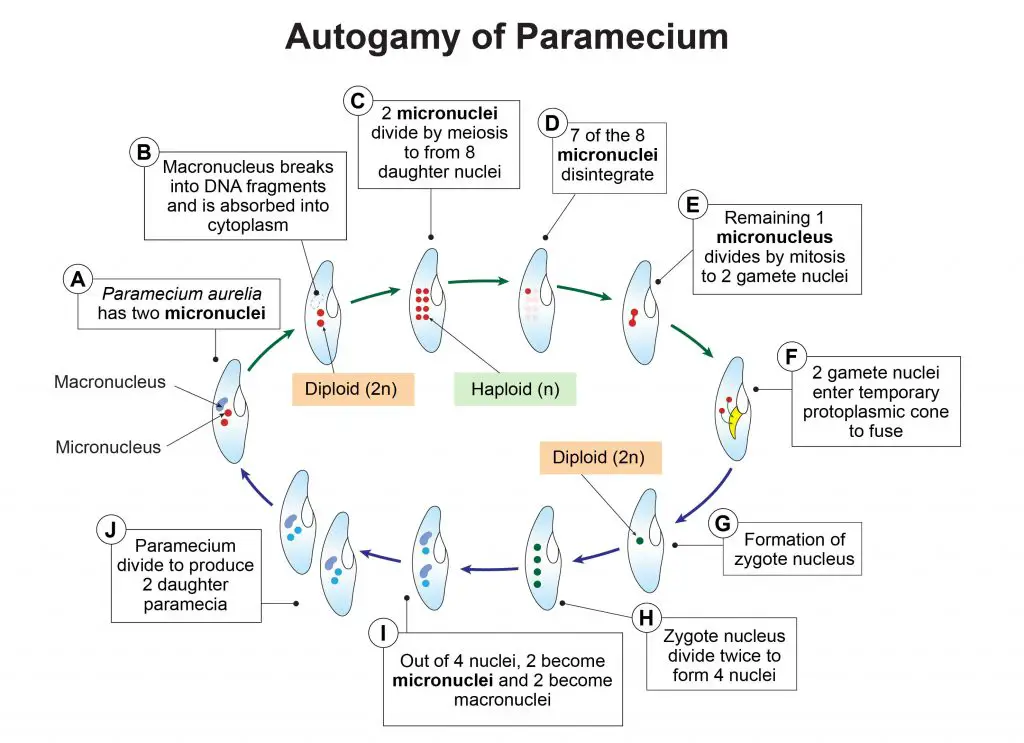
[In this figure] Steps of autogamy of Paramecium aurelia.
(A) P. aurelia consists of one macronucleus and two micronuclei.
(B) Macronucleus grows in size and breaks into DNA fragments which are absorbed by the cytoplasm.
(C) Two diploid micronuclei divide by meiosis to produce 8 haploid daughter nuclei.
(D) Out of these 8 micronuclei, 7 disintegrate.
(E) The remaining one nucleus divides by mitosis and produces two nuclei, now called gamete nuclei.
(F) A temporary protoplasmic cone develops near the mouth. The two gamete nuclei enter this cone.
(G) A fusion of two gamete nuclei produces a diploid nucleus or synkaryon. This zygote nucleus contains all genes in homozygous condition.
(H) Zygote nucleus divides twice by mitosis to produce four nuclei.
(I) Two of these nuclei grow and become two macronuclei and the remaining two become micronuclei.
(J) Each micronucleus and the body of paramecium now divide and produce two daughter paramecia, each with a new macronucleus and two micronuclei.
Endomixis and Cytogamy
Endomixis and cytogamy are two less common methods of reproduction in paramecium. In brief, endomixis happens in a single Paramecium aurelia cell to create nuclear reorganization and rejuvenates its macronucleus. There is no meiosis and no nuclear fusion in this process.
In cytogamy, two paramecia form a pair in a way similar to conjugation. However, there is no nuclear exchange between individuals. Two haploid gamete nuclei in each cell fuse within the original cell. The individuals now separate and divide. Also, a new macronucleus is formed like in conjugation. If you are interested in the detail of endomixis and cytogamy, check out https://www.studyandscore.com/studymaterial-detail/paramecium-sexual-reproduction-and-asexual-reproduction
Aging – Does paramecium get older?
Yes, paramecium will get older, but not at the level of individuals. It is more precise to describe it as the aging of the population.
Aging (or called clonal aging) happens when a group of paramecia grow by asexual reproduction (which cell divisions occur by mitosis rather than meiosis) for a long period of time. Aging leads to a gradual loss of vitality. In some species, such as the well-studied Paramecium tetraurelia, asexual reproduction only allows the paramecia to undergo about 200 fissions. After that, if cells fail to perform autogamy or conjugation, these aged paramecia stop their growth and die.
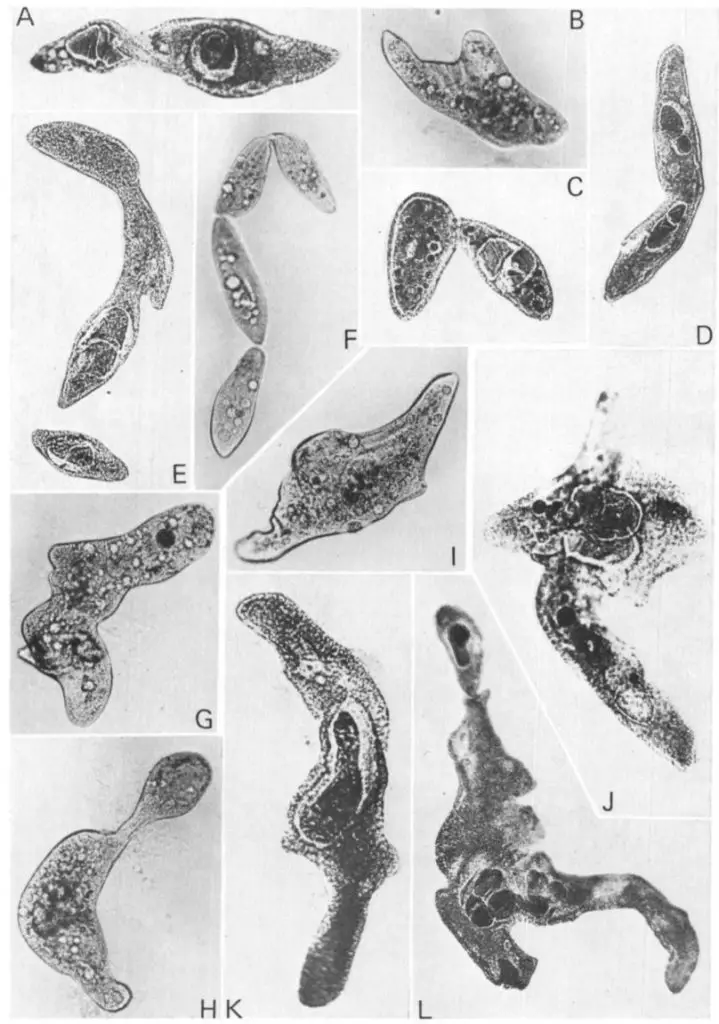
[In this figure] Examples of abnormal cells appeared in old paramecium cultures.
https://jcs.biologists.org/content/41/1/177
The role of macronuclei in aging
We have learned that autogamy and conjugation can rejuvenate paramecium’s macronuclei, does the disability of macronuclei lead to paramecium’s aging?
In fact, scientists proposed a similar hypothesis. In 1986, Karl J. Aufderheide (Texas A&M University) performed a transplantation experiment (through an injection under the microscope) in paramecia. When the macronuclei of young paramecia were injected into paramecia of standard clonal age, the lifespan (how many divisions are allowed) of the recipient was prolonged. In contrast, the transfer of cytoplasm from young paramecia did not prolong the lifespan of the recipient. These experiments beautifully showed that the macronucleus, rather than the cytoplasm, is responsible for clonal aging.
More studies demonstrated that DNA damages accumulated in the macronucleus appear to be the cause of aging in paramecia. Nuclear rearrangement by autogamy or conjugation can reset these DNA damages, resulting in the rejuvenation of paramecium cells. These studies established the “DNA damage theory of aging”, which helps us a better understanding of aging in humans.
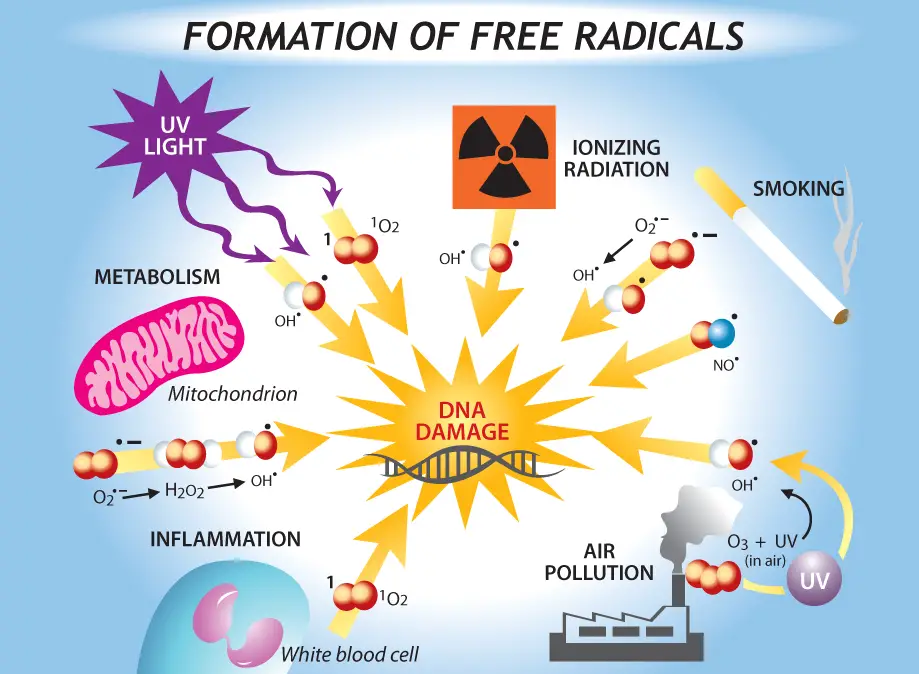
[In this figure] DNA damage is the key of aging.
Many biomedical researchers are working on the prevention and repair of DNA damages to prolong human lifespan.
Photo credit: https://sphweb.bumc.bu.edu/otlt/mph-modules/ph/aging/aging3.html
Paramecia are also good landlords – Endosymbiotic relationship and Endosymbionts
A heterotroph is an organism that cannot produce its own food, instead, taking nutrition from other sources. Since paramecium feeds on other microorganisms to obtain energy, paramecium is a heterotroph.
However, some species of paramecium (for example, Paramecium bursaria and Paramecium chlorelligerum) allow green algae (called Zoochlorella or Chlorella) to live inside its cytoplasm and provide the paramecium cell (the host) with nutrients produced by photosynthesis. At the same time, paramecium provides the algae with movement and protection, as well as carbon dioxide and nitrogen components that are needed for photosynthesis. This mutualistic relationship between paramecium and algae calls “endosymbiotic relationship”.
[In this video] Paramecium bursaria. 40x-600x. Darkfield, ploarized light.
Paramecium bursaria is a species of paramecium that has a mutualistic symbiotic relationship with green algae.
How endosymbiotic relationship start
The endosymbiotic relationship initiates when the P. bursaria cell swallows the green algae by phagocytosis. Rather than digesting, the host paramecium stores the symbiotic algae in vacuoles as endosymbionts. When the paramecium moves towards areas of greater light intensity, algal photosynthesis supplies each partner with photosynthetic nutrients.
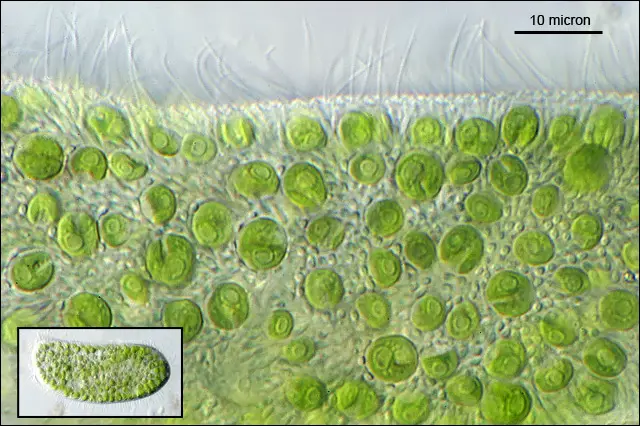
[In this figure] A closer look at the symbiotic algae, Chlorella, that gives it it’s green color.
Photo credit: Charles Krebs.
The endosymbiotic relationship between paramecium and algae is facultative, but not obligate mutualism. P. bursaria and Zoochlorella can survive without the others. However, algae-free P. bursaria are rare in nature. P. bursaria growth is enhanced in cells harboring algal symbionts compared to algae-free cells.
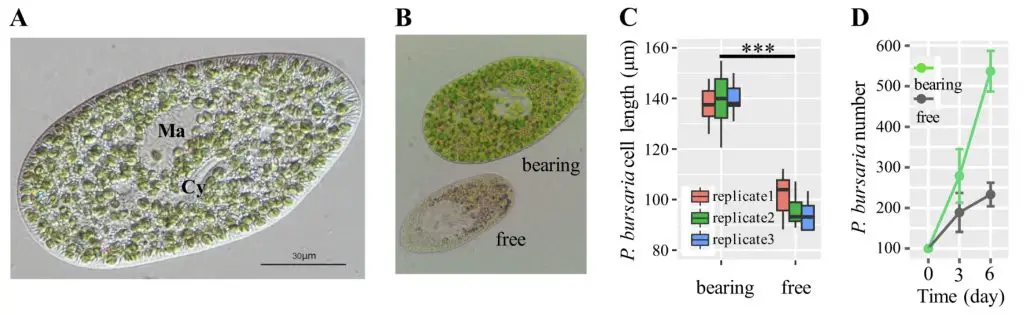
[In this figure] A study of the endosymbiotic relationship between P. bursaria and chlorella.
Scientists compared the difference between algae-bearing and algae-free P. bursaria. (A) Microscope image of a typical P. bursaria cell. “Ma” is macronucleus; “Cy” is cytopharynx. (B) Microscope images of algae-bearing and algae-free P. bursaria. (C) Algae-bearing cells are larger and longer than algae-free P. bursaria. (D) Algae-bearing paramecia grow faster than algae-free cells. The original P. bursaria cell number is 100. The experiments were done in 6 days and fed with bacteria E. coli.
Source: “Genetic basis for the establishment of endosymbiosis in Paramecium” The ISME Journal volume 13, pages1360–1369(2019). https://www.nature.com/articles/s41396-018-0341-4
The benefit of endosymbiotic relationship
Interestingly, endosymbiotic algae also protect their host paramecia from predators. One of the well-studied predators of paramecia, Didinium nasutum, tends to keep away from P. bursaria hosting endosymbiotic green algae. D. nasutum prefers P. caudatum, or P. multi-micronucleatum which don’t have endosymbiotic partners. Scientists hypothesize that the endosymbiotic green algae within P. bursaria discourage predation by D. nasutum by releasing distasteful metabolites that repel them.
Much like roommates adapting to each other’s schedule, the host paramecium and endosymbiotic algae have good communication and are able to synchronize with regard to the timing of cell division and growth. Endosymbiotic algae can even adjust photosynthesis according to the circadian rhythms in the paramecium host.
Other endosymbionts
Bacterial endosymbionts (mostly Gram-negative bacteria) are also found in species of paramecium. It is not known whether in nature this relationship is mutually beneficial or not. Some intracellular bacteria, known as Kappa particles, give paramecia the ability to kill other strains of paramecium that lack Kappa. Killing is mediated by a poison (paramecin) secreted from Kappa particles. Paramecium strains possessing these particles are known as “killer paramecia” and are immune to the poison. Endosymbiotic Kappa particles are inheritable during reproduction.
How does a paramecium move?
As we mentioned in our blog post “the structure of paramecium cell“, paramecium cell moves and collects food using the tiny hair-like structures called cilia. These cilia are in constant motion and help paramecium move around. Paramecium can swim as fast as four times its body length per second. Paramecium can also change its direction by beating the cilia in a reverse way.
[In this video] The movement of Paramecium caudatum under a microscope.
How does a paramecium eat?
Paramecia eat microorganisms that are smaller than them, like bacteria, algae, and yeasts. A single paramecium has the ability to eat 5,000 bacteria a day. As the paramecium moves forward, rotating around its own axis, food materials (like bacteria and algae) flow closer to the opening of the oral groove. To gather the food, the paramecium uses its oral cilia lining the oral groove to sweep the food along with some water into its cell mouth. The food goes through the cell mouth (cytostome) into the gullet (cytopharynx). Through a process known as phagocytosis, the food is packed into food vacuoles for digestion.
The food vacuoles then fuse with lysosomes, and the enzymes enter the vacuole to digest the food contents. The nutrient molecules from digested food then go into the cytoplasm and the vacuole gets smaller and smaller. Once the vacuole reaches the anal pore with all of its digested nutrients, it ruptures and spills all of its waste material into the environment.
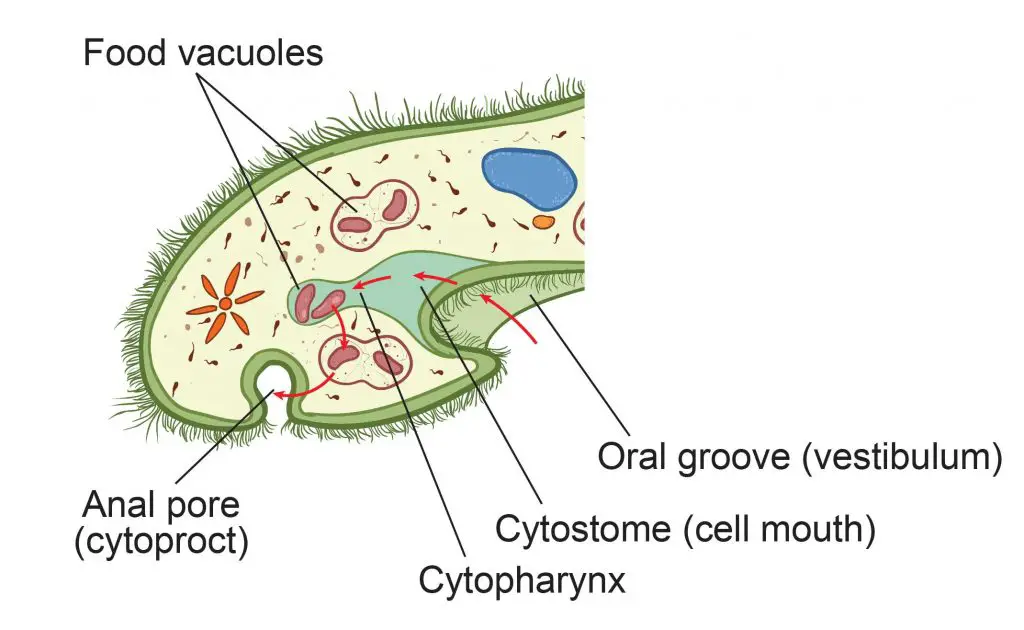
[In this figure] The feeding system of a paramecium.
The red arrows indicate the process of feeding and digestion.
[In this video] How a paramecium eats.
[In this video] Paramecium feeding
An amazing microscopic HD video showing the detachment of food vacuoles at the end of cytopharynx. You can see the movement of oral cilia pushing the food forward.
[In this video] A paramecium takes a poop.
At the time 1:13, you will see a big food vacuole on the left bottom is ready to drop!
How does a paramecium breathe?
Because paramecium is a single-celled organism, oxygen and carbon dioxide can freely diffuse in and out of the cell body. The water flux into the oral groove also increases the efficiency of gas exchange.
Does a paramecium have cytoplasmic streaming?
Yes, cytoplasmic streaming, also called protoplasmic streaming or cyclosis, plays an important role in cell processes since it promotes the movement of the fluid substance (cytoplasm). Cytoplasmic streaming is common in plant cells and large single-cells animal cells.
The function of cytoplasmic streaming
Paramecium is a pretty large cell and relies on cytoplasmic streaming to exchange nutrients and metabolites between the cytoplasm and organelles. Cytoplasmic streaming circulates the cytoplasm and organelles around the paramecium cell. For example, the food vacuoles move around via cytoplasmic streaming to distribute the nutrients in the cell.
The mechanism of cytoplasmic streaming
Cytoplasmic streaming is generated by the movement of motor proteins “myosin” on the cytoskeleton “actin” filaments. As the myosin molecules “walk” along the actin filaments dragging the organelles with them, the circulation of cytoplasmic fluid starts.
[In this video] The cytoplasmic streaming in Paramecium bursaria carries its algal endosymbionts circulating around the cell.
What senses do paramecium have?
The paramecium cannot see, taste, or hear. However, there is evidence suggesting that paramecia have some sense of “smelling” and can respond to certain chemical cues (like glutamate) in their environment.
Paramecia can also sense the intensity of light and prefer to stay in an environment of dim light (because of their food, bacteria, and yeasts, like this environment, too). P. bursaria may move toward bright light to favor the photosynthesis of endosymbiotic green algae. Paramecia also sense pH value and temperature, too. The slightly acidic environment is usually associated with the overgrowth of bacteria.
Paramecium’s cilia can sense its surrounding by touching. This can guide the paramecium to turn around when it bumps into something or fire its trichocytes when it encounters predators. However, the sensor may not be very sensitive, so we frequently see an amoeba slowly approaching and engrafting a paramecium unconsciously.
[In this video] Amoeba hunts and eats paramecia.
Does a paramecium have learning and memory?
Learning is a fundamental process in nervous systems. However, microorganisms without a nervous system have been shown to have simple learning abilities. The question of whether paramecia exhibit learning has been a topic of great scientific interest. However, it is not yet conclusive. Specifically, in 2006, a group of scientists reported that Paramecium caudatum can be trained to associate lighting conditions and electric shocks in its swimming medium. Scientists proposed, based on this observation, that microorganisms with no nervous system may save their cell memory by the modification of DNA (a concept called “epigenetic learning”).
However, another study in 2017 suggested that paramecia can only learn to associate the bright side of its swimming medium with electric current and not the dark side. The whole picture of learning and memory could be much more complicated than we think. Looking forward to hearing more new studies on this very interesting topic.
Can paramecia communicate?
Paramecia may talk to their neighbors by releasing chemicals and cilia touching. Other than that, research published in 2009 reported a very interesting observation – “Paramecia may communicate through light!”
You can read this article for free on the website of PLoS One. Daniel Fels documented the interactions among different populations of Paramecium caudatum, separated by glass. Even through glass, the cells affected cell division and energy uptake in neighboring cell populations. Because the glass barriers effectively prohibit the transfer of chemical signals, Fels infers that these simple organisms are using a form of weak electromagnetic radiation, so-called biophotons, to communicate. Pretty amazing, right?
References
Paramecium: Sexual Reproduction and Asexual Reproduction
“Clonal aging in Paramecium tetraurelia. II. Evidence of functional changes in the macronucleus with age” Mechanisms of Ageing and Development. Volume 37, Issue 3, January 1986–January 1987, Pages 265-279
“Clonal death associated with the number of fissions in Paramecium caudatum” Y. Takagi, M. Yoshida. Journal of Cell Science 1980 41: 177-191
“Feeding Behaviour of Didinium nasutum on Paramecium bursaria with Normal or Apochlorotic Zoochlorellae”
“Diversity and Evolution of Algae: Primary Endosymbiosis.” De Clerck, O., Bogaert, K., Leliaert, F. 2012. Advances in Botanical Research 64, 55–86. ISSN 0065-2296.
“Genetic basis for the establishment of endosymbiosis in Paramecium” The ISME Journal volume 13, pages1360–1369(2019).
“Chemosensory Signal Transduction in Paramecium” Judith L. Van Houten, Wan Qing Yang, Adam Bergeron. The Journal of Nutrition, Volume 130, Issue 4, April 2000, Pages 946S–949S,
“Discrimination Learning in Paramecia (P. caudatum)”. Armus, Harvard L.; Montgomery, Amber R.; Jellison, Jenny L..The Psychological Record. (2006) 56 (4): 489–498.
“Epigenetic learning in non-neural organisms”. Ginsburg, Simona; Jablonka, Eva Journal of Biosciences. (2009). 34 (4): 633–646.
“Paramecium Learning: New Insights and Modifications” Abolfazl Alipour, Mohammadreza Dorvash, Yasaman Yeganeh, Gholamreza Hatam. BioRxiv. Posted November 29, 2017
“Cellular Communication through Light” Daniel Fels. PLoS One. 2009;4(4):e5086. doi: 10.1371/journal.pone.0005086.
Related posts
The Biological Classification of Paramecium – Name, History, and Evolution
The Structure of Paramecium Cell
Pingback: What does Paramecium eat? – Dr. Biology Questions and Answers
Pingback: How does Paramecium eat? – Dr. Biology Questions and Answers